|
August 26, 2002
|
|
 |
 |
| Structural biology of DNA repair, part 2 | ||
| Contact: Paul Preuss, paul_preuss@lbl.gov | ||
|
|
The ties that bind The binding head is compact, but each of its Rad50s has a long tail, extending up to 600 angstroms in mammals, which consists of a coil of amino acids coiled back around itself -- like a twisted fiber pinched in the middle, its two halves wound around each other to form a loose piece of twine.
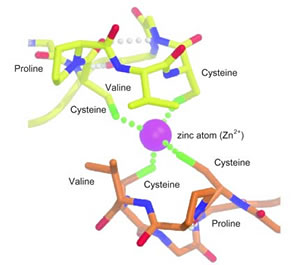
At the pinch in the middle of the primary coil, where the tail bends back on itself, there is a sequence of four amino acid residues similar in all the organisms studied. The first and fourth residues in this sequence are always cysteines, with various other residues in the second and third places -- thus the label "CXXC motif." Through x-ray crystallography the researchers learned that the CXXC motifs were shaped like hooks, with the potential to grapple the tails of other Rad50 proteins. Using laboratory techniques of gel filtration and ultracentrifugation, the group confirmed that successful linkage of two Rad50 proteins depends on the presence of zinc. Chemical and structural studies showed that four cysteines (two each, in a pair of Rad50 tails) bind to a single, doubly ionized zinc atom. Discovery of this zinc-hook mechanism immediately suggested ways the Mre11 complex could connect DNA strands and bring them together. A biological multitool kit In one micrograph, the twin tails of a single binding head link to each other, forming what looks like a finger ring, with the binding head as its stone; an even more unusual micrograph shows a single-headed complex of this kind actually bound to a length of DNA. These conformations suggest distinct mechanisms by which linked Mre11 complexes can bridge sister chromatids or perform nonhomologous end-joining, and how even a circular, single-headed Mre11 could bring together two broken DNA ends.
"Identification of the Rad50 zinc hook will allow us to understand the mechanism for dynamic assembly and disassembly of the Mre11 complex," says Tainer, "which is critical for the repair of double-strand breaks in DNA and thus for the avoidance of cancer-causing mutations." The discovery is exciting not only for the questions it answers but for the new questions it raises. Do binding heads begin their work separately, thrashing their tails about until they grapple another bound complex that's likely to help complete the repair? Do they link tails first and search together for broken DNA to repair? Other models are possible; which are correct and how the process works in detail are yet to be determined, but other coordinating proteins and interactions are likely to be involved. Thus finding the correct model requires the complete NCI-funded team. SBDR's leaders emphasize the cyclical nature of their work, first drawing on genome sequencing and biochemical and genetic studies to suggest the proteins most worth studying, then determining the structures of these proteins through x-ray crystallography and other methods, in search of understanding of how they work together to accomplish complex biological tasks. This knowledge, in turn, suggests biological experiments that can give clues to the make-up and mechanisms underlying the complex machines that mediate DNA repair, filling in more details of their intricate interactions -- a new kind of research design cycle that provides a novel prototype for productively linking structure and chemistry with biology. Additional information: |
||||||||||||||||||