February 16, 1999 |
|
 |
 |
 |
BERKELEY, CA — In a few seconds an
origami artist can fold a sheet of paper into a bird or flower or pagoda or other
intricate shape. In much less time a string of amino acids can fold itself into a protein,
the kind of molecule that comes in many thousands of complex shapes and does most of the
work of life. Origami can be taught, but no one knows how proteins fold themselves so
quickly into the same shapes virtually every time.
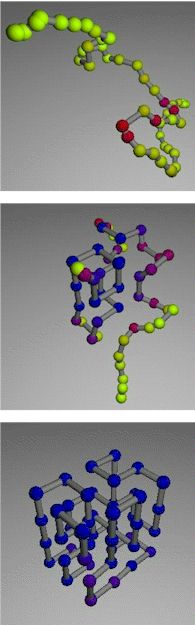
Now, computer models devised by Daniel Rokhsar and his colleague Vijay Pande of the Department of Energy's Lawrence Berkeley National Laboratory, working at the National Energy Research Scientific Computing Center (NERSC), have revealed unexpected regularities in the pathways of protein-like structures. They report their findings in the Proceedings of the National Academy of Sciences, February 16, 1999 (vol. 96, no. 4). "We're interested in the physical mechanisms by which biomolecules achieve their structures," says Rokhsar, who is head of the Computational and Theoretical Biology Department in Berkeley Lab's Physical Biosciences Division and a professor of physics at the University of California at Berkeley. The precise structure of biomolecules often reveals their functional secrets, a fact that became clear in the early 1950s when Linus Pauling solved the alpha-helix structure of keratin protein and Watson and Crick solved the double-helix structure of DNA. Protein shapes determine everything from the texture of hair and horn to the catalytic coupling and uncoupling of innumerable enzymes essential to keep life's processes humming. Misfolded proteins can cause disease; in humans, sickle-cell disease and other anemias, for example, are caused by the misfolding of hemoglobin, whose normal structure, resembling a miniature spring-clip, allows it to capture, transport, and release oxygen in the bloodstream. For any protein there is a "native state conformation," a thermodynamically most-stable structure that depends upon the energy of the bonds that form when the acid residues come close together, and upon whether a given group of amino acid residues is hydrophobic or hydrophilic, and so on. But newly manufactured proteins are far from their native state. Proteins are punched out rather like ticker-tape by ribosomes that add amino acids one at a time. Although the order of the amino acids is ultimately specified by a length of DNA (the gene for that protein), how the order specifies the protein's distinctively folded structure and directs pathways to that structure is not yet understood. "Protein chains all fold differently—even proteins of the same kind fold into their final state by sampling many different conformations—because they start from different initial states," says Rokhsar. "Yet somehow they start from an unfolded state and achieve the folded structure quickly, reliably, and reversibly." To demonstrate the magnitude of the challenge, Rokhsar suggests contemplating a single node—a stand-in for a single amino acid residue—represented by a ball on the end of a stick. "Let's limit to five the directions the next stick-and-ball can extend—right, left, up, down, or straight ahead," says Rokhsar. "If there are five links in the chain, that's five to the fifth ways the chain could fold, 3,125 possibilities. If there are a hundred links in the chain—not unusual for a protein—there would be something like 10 to the 30th possibilities. If you tried them randomly, even a trillion times a second, it would take longer than the age of the universe to get the right structure." Rokhsar and Pande, who is a Miller Postdoctoral Fellow at UC Berkeley's Department of Physics, approached the problem by designing a protein-like model heteropolymer of 48 units whose properties define a stable "native structure"—a compact lattice in three dimensions with each bend at a right angle, resembling a jungle gym made of Tinker Toys. Using the Cray T3E computer at NERSC, Rokhsar and Pande repeatedly unfolded the model by raising its (simulated) temperature, then lowered the temperature and watched it refold itself. For each folding sequence they separately tracked the position of each of the 48 "mers," the units equivalent to a protein's amino acid residues. Even with a model far less complex than most real proteins, the number of possible initial conformations is astronomically large, and each path to stability is virtually unique. By sampling the state of the writhing polymer every 6,000 iterations—taking a single-frame snapshot of the shape—the researchers made movies that showed the model polymer seeking and eventually finding its stable state. Typically some three-quarters of a million iterations were required before the model polymer stabilized. The average position change of each unit was recorded from frame to frame, and the rate of change was color-coded—from yellow for units that thrashed continually, through the spectrum to blue for those that held still, at least temporarily. This data could be arranged in "fluctuation smears" to give a cumulative picture of the position of the units at any moment in the process. Remarkably, Rokhsar and Pande discovered common features among the numerous folding pathways. At first the unfolded polymers fluctuated wildly through several hundred thousand configurations—then suddenly settled into a partially folded intermediate state, in which a stable core structure was accompanied by flailing loops and dangling ends. After another couple of hundred thousand iterations, the polymer abruptly locked into its native state. These sudden transitions are evocative of phase changes, like the changes from a gas to a liquid to a solid. There are distinct classes of intermediate states for the model polymer, however, which correspond to different groups of units that temporarily achieve stability during the intermediate phase. Each class of intermediate states represents a set of related pathways from the unfolded to the native state. When Rokhsar and Pande repeated their simulations with model polymers of 64 and 80 mers, the folding pathways also grouped themselves into separate classes of intermediate states. These intermediate phases are closely analogous to partially unfolded states (PUFs) which have been observed in real proteins, as well as to intermediate states inferred to exist in other real proteins. It is likely that knowledge of PUFs, plus inferences about similar phases from other protein studies, can predict transition states of some kinds of proteins in the real world. Rokhsar's and Pande's discovery of well-defined transition states in model-polymer folding has important implications for the development of a general theory of protein folding. Verifying these results using models with atomistic detail is the next important step. The lattice model of protein folding can be seen in spectacular action by visiting http://hubbell.berkeley.edu/nsb.html. Also see the Pande Group website and the image gallery on that site. The Berkeley Lab is a U.S. Department of Energy national laboratory located in Berkeley, California. It conducts unclassified scientific research and is managed by the University of California. |